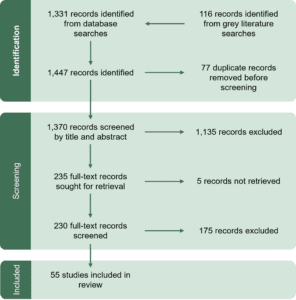
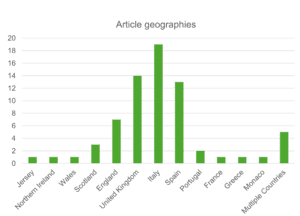
5.1
This review highlights that targeted scientific evidence from the last 25 years, which quantifies the effects of MPAs on the extent and condition of benthic habitats and their associated communities, is limited to relatively few studies. While MPAs are widely promoted as a tool for preserving and restoring marine ecosystems, much of the existing research has focused primarily on mobile or commercially valuable taxa, particularly fish. Indeed, 20% of the studies identified in this review are focused exclusively on fish assemblages associated with benthic habitats, with comparatively less attention paid to benthic systems such as seafloor invertebrate communities, macroalgae, and structural habitats like corals, sponges, or seagrasses. This taxonomic and thematic bias continues to constrain our understanding of how MPAs affect benthic habitats and their associated communities, especially those structured by slow-growing, habitat-forming organisms, such as maerl and horse mussels (Modiolus modiolus).
5.2
Nevertheless, most of the studies reviewed reported predominately positive ecological responses from benthic habitats and their associated biological communities, attributed to the implementation of MPAs. These benefits are reflected in multiple metrics, including increased species richness, shifts in community composition,[5-7] enhanced biomass of benthic organisms,[8-12] and improvements in habitat complexity,[6] alongside evidence of enhanced recovery from natural disturbance.[13] Although the evidence base remains limited, these findings collectively underscore the potential for MPAs to contribute meaningfully to the conservation and recovery of benthic ecosystems when appropriate conditions and management frameworks are in place.
Benthic community responses to protection
5.3
Changes in community composition, species richness, abundance, and biomass are among the most commonly reported ecological responses to MPAs. However, these changes are not uniform across all habitats or ecological contexts, and often vary depending on habitat type, assemblage structure, species-specific ecological traits, or a combination of these factors. Nevertheless, a recurring pattern observed across the literature is that MPAs tend to promote shifts in the structure of benthic communities, with assemblages in protected areas diverging over time from those in adjacent areas without protection. The direction and magnitude of these changes, however, can vary considerably, with some communities showing pronounced recovery [6, 10] and restructuring while others remain stable and exhibit minimal change.[14]
5.4
Among the most consistent ecological responses to protection is the transformation of benthic community composition. Several studies reviewed reported an increase in biodiversity and species richness within MPAs in comparison to their unprotected counterparts,[5, 15-17] particularly among sessile and slow-growing organisms, such as sponges, corals and bryozoans. These changes are often characterised by a shift away from disturbance-tolerant or opportunistic species toward more structurally complex and ecologically functional assemblages [5, 15], indicating a broader move towards healthier and more stable benthic ecosystems.
5.5
The benthic community changes observed within the Lyme Bay MPA, located off the south coast of England, provide a compelling example of how spatial protection can influence benthic community composition over time.[6] Between 2008 and 2011, the structure of benthic assemblages shifted significantly, marked by substantial increases in the abundance of several habitat-forming and ecologically important taxa. The bryozoan Pentapora fascialis, a colony-forming invertebrate, increased by an average of 385%, while branching sponges rose by 414%. Hydroid abundance increased by 229%, although high spatial variability limited statistical confidence. Cold-water corals Parerythropodium mammillatum and Alcyonium digitatum exhibited increases of 467% and 2,541%, respectively. Meanwhile, the sea fan Eunicella verrucosa, a key structural species, showed a 636% rise in mean abundance.[6]
5.6
These patterns suggest the development of more mature and structurally diverse benthic communities, showing greater structural complexity and dominance of slower growing taxa over early colonising species. In turn, these compositional changes reflect improved habitat quality and enhanced ecological functioning, consistent with broader patterns of benthic recovery documented in effectively managed MPAs. Such findings highlight the potential for MPAs to enable community-level recovery following anthropogenic disturbance, especially where damaging activities like bottom-contact fishing have historically degraded habitat quality. Through protection, MPAs can allow complex assemblages to re-establish over time, thereby enhancing ecosystem resilience, biodiversity, and habitat integrity.
5.7
Beyond changes in benthic community composition, numerous studies have documented increases in the biomass of marine species within MPAs [9, 12, 18], particularly among commercially targeted taxa such as fish and scallops.[19, 20] These increases are often attributed to the cessation or reduction of extractive activities under MPA protection, allowing populations to recover from prior exploitation. However, the response to protection is not universal, and outcomes can vary depending on a species’ trophic role and its interactions with other species within the community, as well as the broader ecological context of the protected area.
5.8
Within commercially exploited species, there are consistent trends of increases in biomass, abundance, and body size within MPAs. For example, in the South Arran Marine Reserve (now designated as a NCMPA), scallop populations of Aequipecten opercularis and Pecten maximus were, on average, significantly larger and exhibited greater exploitable and reproductive biomass than those found outside the MPA.[20] Moreover, densities of these species were substantially higher within the reserve compared to adjacent, unprotected areas, with densities declining progressively with increasing distance from the MPA boundary, suggesting a potential spillover effect into surrounding fished areas.[19]
5.9
Similarly, commercially targeted sparid fish such as Diplodus sargus and Diplodus vulgaris have shown significant increases in both size and biomass within protected areas that restrict all fishing compared to adjacent fished sites, a trend that has been documented across several studies.[9, 12, 21, 22] However, it is important to note that increased biomass within MPAs can often reflect the presence of larger individuals rather than a higher density or abundance of individuals.[11] In these cases, the absence of fishing pressure enables fish to survive longer and grow into larger size classes, which ultimately enhances reproductive output and supports long-term population sustainability, even if overall abundance increases are not observed in short terms (~3-5 year) studies. These findings also reflect a broader bias in the literature, where monitoring and assessment efforts have disproportionately focused on commercially valuable species, potentially overlooking responses among other benthic species that may not exhibit such conspicuous or economically relevant changes.
5.10
In contrast, species that are not strongly affected by fishing pressure often show inconsistent or negligible responses to protection within MPAs.[23]. This variability highlights the need to consider species-specific ecological characteristics when evaluating MPA effectiveness. For instance, sedentary or site-attached benthic species with long lifespans and slow maturation rates are more likely to benefit from reduced exploitation and disturbance within MPAs over time. In contrast, highly mobile or opportunistic species may move freely across MPA boundaries or recover quickly due to natural fluctuations, making their response to spatial protection less predictable.
5.11
The broader ecological context of spatial protection – such as habitat quality within the MPA, connectivity to surrounding habitats/populations, and the intensity of fishing pressure in adjacent areas – can significantly influence ecological outcomes. Importantly, the demographic changes induced by protection frequently extend beyond individual species, driving shifts in trophic interactions and broader community dynamics. As populations of predators recover within MPAs, they can exert increased top-down control on lower trophic levels, altering species abundances and interactions across the food web.
5.12
For example, in the Kattegat no take zone (NTZ), the recovery of predatory benthivore fish has been associated with notable declines in brittle star populations, specifically Amphiura filiformis and Amphiura chiajei, likely driven by increased predation pressure.[5] This negative correlation between benthivore fish biomass and brittle star biomass was observed within the NTZ, in contrast to the absence of such a relationship in adjacent trawled reference areas. These patterns are indicative of trophic restructuring, whereby the increase in benthivore flatfish following the cessation of fishing within the NTZ has likely contributed to the suppression of dominant brittle star populations through enhanced top-down control.
5.13
Similar patterns have been observed in sea urchin populations, which frequently decline within MPAs as predatory fish recover and recover their ecological roles.[10] As predator assemblages recover within MPAs, increased predation pressure can significantly reshape the size-frequency distribution of sea urchin populations, often leading to a notable reduction in intermediate size classes. This pattern reflects size-selective predation and provides strong evidence of top-down control within benthic food webs.
5.14
In the case of the sea urchin species Paracentrotus lividus, predation risk was found to correlate positively with protection level and the biomass of predatory fish within a fully protected area.[22] Elevated predator densities within fully protected zones appeared to offset the expected positive demographic response of sea urchins to reduced harvesting. The resulting population structure was skewed, with a dominance of small and large individuals and a marked deficiency of medium-sized urchins. This bimodal distribution likely arose from the ability of juveniles to seek refuge in crevices and the reduced vulnerability of larger individuals due to predator handling limitations, thereby concentrating predation pressure on intermediate size classes. This demographic shift led to a reduction in overall grazing pressure, which in turn facilitated an increase in benthic primary producers. Within the MPA, this was reflected in a higher cover of turf-forming and erect branched algal species compared to adjacent unprotected areas.[22] This highlights the cascading ecological effects of predator recovery and the restoration of natural trophic interactions.
5.15
While such cascading ecological effects are often attributed to protection, it is also essential to recognise that natural ecological variability can sometimes mask or confound these patterns. For example, great scallops exhibited a marked increase in juvenile abundance within protected areas, with densities substantially higher than those in adjacent, unprotected areas[19, 20] While this trend aligns with expectations under reduced fishing pressure, it is important to note that the overall abundance of juvenile scallops in the region has been observed to fluctuate on a roughly biennial basis, alternating between low and high recruitment years.[19] Such natural variability in recruitment cycles complicates the attribution of observed changes solely to MPA protection. Short-term assessments that coincide with unusually high or low recruitment periods may overestimate or underestimate the true effects of protection. This highlights the critical importance of long-term monitoring and the inclusion of appropriate reference sites to disentangle protection effects from background ecological dynamics, ensuring that observed changes are interpreted within the correct temporal and environmental context.
5.16
In summary, the ecological responses of benthic species and communities to MPAs are diverse, complex, and strongly context dependent. While consistent trends such as increases in biomass, body size, and biodiversity are well documented, particularly among commercially targeted and structurally important benthic taxa, these outcomes are shaped by a range of interacting factors. Habitat type, assemblage composition, species-specific life-history traits, and broader ecological conditions all influence the direction and magnitude of community change. Importantly, MPAs can facilitate trophic restructuring through the recovery of predator populations, leading to cascading effects that alter benthic community composition, suppress grazer populations, and promote the proliferation of habitat-forming species. However, natural ecological variability, such as episodic recruitment and environmental fluctuations, can obscure or confound these effects, highlighting the importance of long-term, spatially replicated monitoring. A nuanced, ecosystem-based approach to MPA evaluation that accounts for both protection effects and background ecological dynamics is therefore essential for accurately assessing outcomes and guiding effective conservation management.
Recovery of benthic habitats within MPAs
5.17
The recovery of benthic habitats represents a key ecological outcome of effective implementation of MPAs. Beyond changes in species composition and biomass, improvements in habitat structure and integrity are critical indicators of ecosystem restoration. Within MPAs, the exclusion or reduction of damaging activities, particularly bottom-contact fishing, allows physically degraded habitats to begin a natural recovery process, often marked by increased complexity, stability, and biological productivity.
5.18
Structurally complex habitats such as biogenic reefs, sponge grounds, seagrass meadows, and maerl beds have shown strong potential for recovery under sustained protection. As previously discussed in paragraph 5.5, the cessation of mobile fishing in Lyme Bay MPA has led to the re-establishment of diverse epifaunal assemblages and the return of key habitat-forming species, including branching sponges and cold-water corals.[6] These organisms play a foundational role in creating three-dimension benthic habitat with vertical surfaces and increased surface area that provide shelter and substrate for a variety of associated fauna.
5.19
The trajectory, pace, and outcomes of benthic habitat recovery are influenced by multiple factors, including substrate type, the intensity and duration of historical disturbance, and the regenerative capacity of the species. Maerl beds, formed by slow-growing calcareous red algae, are particularly sensitive to physical disturbance, and their slow rates of recovery are widely recognised. Consistent with this, evidence suggests that maerl beds can exhibit marked differences in morphology and structural complexity depending on the duration of protection. For instance, maerl beds within an MPA, which had designated for six years, displayed significantly lower structural development, characterised by smaller size, less spheroidal shape, and lower morphological complexity when compared to those in a long-established MPA that had been designated for 25 years.[24] These findings suggest that while MPAs can promote the recovery of maerl habitats, extended periods of sustained protection are likely required for them to regain their full structural complexity and ecological function.
5.20
Environmental conditions can further influence recovery trajectories. For instance, within the same study, maerl morphology was compared across different depths in the long-established MPA, revealing significant variation. Regardless of growth strategy (ramified versus nucleated), maerl in shallow beds was generally larger, rounder, and more solid, meaning less branched and structurally complex, than maerl found in deeper areas.[24] These patterns underscore the influence of environmental gradients such as hydrodynamics, light availability, and sedimentation rates on habitat structure, suggesting that recovery potential is not uniform even under long-term protection.
5.21
While MPAs can facilitate the recovery of benthic habitats, protection does not always equate to ecological improvement or measurable recovery. In some cases, the absence of observed recovery, or even further degradation, reflects complex underlying pressures or historical impacts that are not easily reversed. A notable example is Strangford Lough, where benthic habitat condition has continued to decline despite protective measures. Between 2003 and 2010, mean densities of horse mussels ( modiolus) in the north basin declined by approximately 83%, from 80 individuals per square metre to just 13.6 individuals per square metre.[7] This represents an average reduction of over 66 individuals per square metre, and a significant deterioration in condition within a relatively short timeframe, despite the area being designated for conservation.
5.22
Such trends highlight the importance of recognising that recovery is not guaranteed simply through spatial protection. In many nearshore environments, particularly those with a long history of anthropogenic disturbance, conservation efforts must contend with shifting baselines. This term refers to the gradual lowering of ecological reference points used in trend assessments over time, as each generation comes to accept a progressively degraded state as normal. The presence of shifting baselines complicates the assessment of MPA outcomes, since current conservation targets may be based on already altered or degraded benchmarks.
5.23
In some instances, habitat recovery may be further constrained by legacy impacts that have pushed ecosystems beyond critical thresholds, significantly reducing their capacity to regenerate without active intervention. This highlights the need for timely protection, ideally implemented before irreversible degradation occurs. It also underscores the potential necessity of active restoration measures, such as reseeding, artificial reef construction, or sediment remediation, particularly in severely impacted areas.
5.24
In addition to historical impacts and environmental conditions, the recovery of benthic habitats can also be influenced by interactions with other species within the community. As previously discussed, trophic dynamics and species-specific responses to protection can shape recovery trajectories in unexpected ways. For example, Laminaria ochroleuca, a habitat-forming kelp species, has been observed to fail to re-establish a canopy forest within an MPA, despite reductions in direct physical disturbance.[25] Instead, persistent grazing by recovering fish populations has limited the survival of juvenile kelp, thereby inhibiting the development of a mature ochroleuca canopy.
5.25
This example highlights how ecological interactions within protected areas can influence habitat recovery outcomes, even in the absence of direct anthropogenic disturbance. It demonstrates that protection alone does not guarantee the re-establishment of all habitat-forming species, particularly when biotic pressures such as herbivory remain high. Such outcomes underscore the complexity of benthic ecosystem dynamics and reinforce the need for holistic, community-level assessments when evaluating MPA effectiveness for achieving seafloor integrity targets. Assessing not only the responses of individual species, but also the interactions among functional groups, is essential for anticipating potential trade-offs and managing ecosystems in a way that supports broader objectives for GES.
Role of disturbance and recovery support
5.26
Both natural and anthropogenic disturbances can significantly alter marine habitats and the composition of benthic assemblages, even within designated MPAs. Extreme weather events can induce rapid and profound alterations in benthic ecosystems, potentially offsetting ecological gains achieved through protection. A well-documented example of this comes from Lyme Bay (United Kingdom), where a severe storm event in 2013 caused widespread disruption to the benthic community.[13] The effects of the storm were more pronounced within the MPA than in adjacent unprotected sites, resulting in greater losses of species and individuals. This disturbance temporarily reversed the protective gains, with community composition within the MPA reverting to a state more closely resembling that of unprotected areas and effectively erasing the structural divergence that had previously developed through protection.
5.27
Despite this setback, post-disturbance monitoring revealed a markedly faster recovery trajectory within the Lyme Bay MPA relative to unprotected sites. Over time, the protected assemblages re-established greater ecological distinctiveness and structural complexity, whereas communities outside the MPA remained in a persistently degraded state. These findings suggest that although the MPA did not confer strong resistance to the acute impact of the storm, it enhanced ecological resilience by facilitating more rapid and complete recovery. This distinction is important as while resistance refers to the ability to withstand disturbance, resilience reflects the capacity to recover. The observed pattern was further supported by evidence showing that recovery from storm damage within the MPA outpaced previous recovery from chronic impacts such as bottom-towed fishing activities.[13] In contrast, benthic assemblages in unprotected areas remained impacted throughout the monitoring period, highlighting the long-term consequences of continued anthropogenic pressure.
5.28
This case illustrates that, although MPAs may not always buffer against the immediate effects of extreme events, they can play a critical role in promoting ecosystem resilience and long-term recovery. Protection appears to enhance recovery capacity by fostering more functionally diverse, structurally complex, and interconnected communities, which are better equipped to rebound following disturbance events.
Effectiveness and limitations of MPAs: implications for management
5.29
MPAs represent a widely used tool for marine conservation and habitat recovery, yet their effectiveness is neither universal nor guaranteed. As this review has shown, MPAs can facilitate the restoration of benthic habitats and communities, enhance biodiversity, and improve ecological resilience. However, these outcomes are contingent on a range of ecological and managerial factors, and protection alone is not always sufficient to ensure recovery and/or progress towards seafloor integrity targets for GES.
5.30
The ecological success of an MPA depends critically on its design, implementation, and enforcement. Key attributes such as size, age, level of protection, spatial configuration, and placement relative to key habitats and ecological corridors collectively influence the extent to which an MPA can mitigate anthropogenic impacts and support ecological recovery. Older, well-enforced MPAs are consistently associated with more pronounced ecological benefits. In contrast, smaller MPAs, those with limited enforcement, or those incorporating partially protected zones (where some human activities remain) tend to show more variable or limited ecological outcomes. Evidence increasingly suggests that fully protected areas, where extractive activities are entirely prohibited across the site, are more effective than partially protected zones in achieving meaningful conservation gains, particularly for benthic habitats and associated communities.
5.31
While MPAs offer localised protection, they are not insulated from broader environmental pressures. Disturbances originating beyond their boundaries, including increased sedimentation, nutrient loading, invasive species, ocean warming, and extreme storm events, can reduce their ecological effectiveness. These external stressors have the potential to degrade habitat condition and alter community structure, even in well-managed protected areas. Nevertheless, as noted in paragraph 5.25, MPAs can increase the resilience of benthic habitats and communities, supporting more rapid and complete recovery following such disturbances. This highlights the importance of integrating MPAs with wider marine management measures that address both local and regional drivers of ecological change.
5.32
Slow ecological response times and historically degraded baseline conditions further complicate assessments of effectiveness. In some cases, benthic habitats may require extended periods to respond to protection due to slow growth rates or limited larval supply. Recovery may also be masked by shifting baselines, where the full extent of historical degradation is poorly understood, and modern reference conditions already reflect diminished ecological states.
5.33
Importantly, the absence of measurable ecological change should not automatically be interpreted as evidence of MPA failure or that spatial protection is an ineffective measure for achieving seafloor integrity targets. In some cases, limited or no apparent response may reflect complex ecological constraints, insufficient time since designation, or misalignment between management measures and the ecological characteristics or recovery timescales of the targeted habitats and communities. In addition, sampling at insufficient time intervals may fail to capture natural seasonal or interannual variability, potentially obscuring underlying trends. Such outcomes do not definitively indicate that an MPA is ineffective but rather highlight the need for careful interpretation and long-term, ecologically informed monitoring. The most effective way to minimise the likelihood of ambiguous or limited outcomes is to carefully plan and design MPAs around the specific ecological characteristics and recovery dynamics of benthic habitats. Aligning spatial protection with habitat-specific needs increases the likelihood of achieving conservation objectives, such as GES, and delivering measurable ecological benefits.
5.34
Given this complexity, robust and habitat-specific monitoring strategies are essential. Benthic communities are often highly variable and tend to respond slowly and non-linearly to management interventions. Effective monitoring must be capable of capturing both annual and seasonal variation across the entire benthic community, including habitat features, sessile and mobile invertebrates, structural taxa, and associated fish assemblages. The reviewed literature demonstrate that where possible, it is critical to gather baseline data prior to MPA designation to establish ecological reference points and enable meaningful assessment of change over time. In addition, monitoring programmes should include sampling at appropriately matched reference sites located in ecologically similar, unprotected areas. This spatial replication allows for better attribution of observed changes to protection effects, helping to distinguish them from broader environmental variation. Long-term, spatially explicit programmes should incorporate benthic indicators, such as habitat extent and structural complexity, to detect ecologically meaningful trends and assess recovery trajectories in systems where responses may be subtle, lagged, or spatially heterogeneous.
5.35
In conclusion, the reviewed literature demonstrates that MPAs can be effective in improving the extent and condition of benthic habitats and their associated assemblages. However, their success depends on careful design, consistent enforcement, and long-term management. Protection measures must be ecologically tailored, adequately resourced, and embedded within broader management strategies that address both local and regional pressures. To restore and sustain healthy benthic ecosystems and their associated assemblages, MPAs must be supported by scientific understanding, adaptive management, and targeted monitoring. Only through this integrated approach can they be full effective for delivering targets for GES under the UKMS.